
Hiện tượng PHS ở ngũ cốc
Lũ lụt là một trong những yếu tố bất lợi phi sinh học có ảnh hưởng nặng nề và toàn diện đến nền sản xuất nông nghiệp, là một trong những nguyên nhân gây ra hiện tượng PHS [1]. PHS được định nghĩa là hiện tượng hạt hoặc phôi bắt đầu phát triển trước khi chúng tách rời khỏi cây mẹ [2]. Đây là hiện tượng rất phổ biến ở các loài thực vật bậc cao, được ghi nhận ở gần 100 loài cây có hoa (thuộc 40 chi và 23 họ khác nhau) [3, 4]. Đặc biệt, PHS xuất hiện trên nhiều loại cây ngũ cốc, điển hình như lúa mì, lúa mạch, ngô và lúa gạo ở hầu hết các khu vực trên thế giới, gây thiệt hại hàng tỷ USD mỗi năm [5] (hình 1).

Hình 1. Hiện tượng PHS xảy ra ở các loại cây ngũ cốc. (A) lúa mỳ, (B) lúa mạch, (C) ngô và (D) lúa gạo.
Ở hầu hết các cây ngũ cốc, cơ chế ngủ nghỉ của hạt (seed dormancy) được chứng minh có liên quan chặt chẽ tới quá trình mất nước xảy ra trong giai đoạn chín của hạt [6]. Khi đó, carbohydrate tích lũy tỷ lệ thuận với lượng nước mất đi theo quá trình phát triển của hạt. Tuy nhiên, các trận mưa trái mùa diễn ra có thể là điều kiện để carbohyrate dự trữ bị "ngậm" nước, từ đó phôi phát triển ngay bên trong vỏ hạt khi còn trên cây (hình 2). Cụ thể, điều kiện độ ẩm cao (do mưa liên tục và kéo dài) kèm theo nhiệt độ cao xâm nhập qua lớp vỏ đã phá vỡ quá trình ngủ nghỉ của hạt ngũ cốc thông qua sự thay đổi của các hormone, đặc biệt là ABA và GA [4, 6, 7].
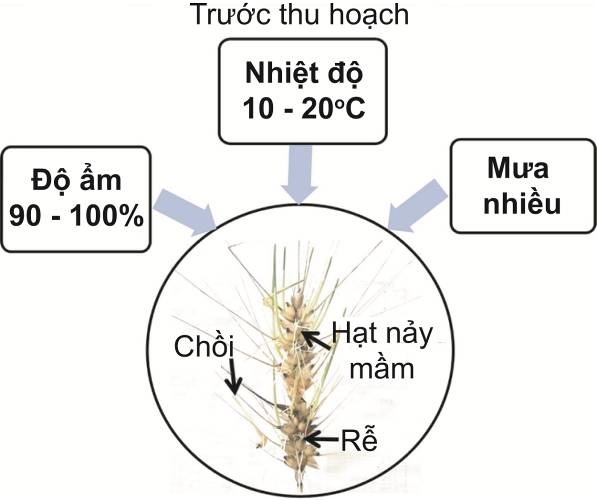
Hình 2. Các điều kiện ngoại cảnh thúc đẩy hiện tượng PHS ở cây ngũ cốc.
Về mặt lý thuyết, ABA là hormone liên quan trực tiếp đến quá trình ngủ nghỉ của hạt, trong khi GA thúc đẩy sự nảy mầm của hạt và phá vỡ trạng thái ngủ đông [8]. Các nghiên cứu trước đây đã chứng minh rằng tỷ lệ ABA/GA điều khiển trực tiếp quá trình chuyển dịch giữa hai trạng thái nảy mầm và phải nghỉ của hạt [4, 6-8]. Trong quá trình phát triển của hạt, việc tổng hợp ABA của cả cây mẹ và trong nội phôi đều ngăn cản PHS xảy ra, vì vậy tỷ lệ ABA/GA luôn được duy trì ở mức cao nhằm duy trì trạng thái ngủ nghỉ của hạt ngũ cốc [9]. Tuy nhiên, kích thích của các bất lợi phi sinh học (mưa nhiều kèm nhiệt độ) đã hoạt hóa biểu hiện của một số gen điều hòa, thúc đẩy quá trình tổng hợp GA, dẫn đến sự mất cân bằng tỷ lệ ABA/GA. Bên cạnh đó, hoạt động của các enzym thủy phân cũng được tăng cường, đặc biệt là α-amlyase (gấp 20-40 lần), từ đó làm bột ngũ cốc trở nên xốp, dính và mất màu, giảm giá trị kinh tế của hạt 20-50% [10]…
Đến nay, cơ chế điều hòa quá trình ngủ nghỉ và nảy mầm ở hạt ngũ cốc thông qua con đường tín hiệu giữa các hormone vẫn chưa hoàn toàn được làm sáng tỏ. Ví dụ, Yan và Chen (2020) [9] đã tóm lược các nghiên cứu trước đây và chỉ ra rằng, ethylene là hormone ít hoạt động trong giai đoạn ngủ nghỉ của hạt, nhưng có xu hướng hoạt động mạnh khi hạt tiếp xúc với nước và bắt đầu nảy mầm. Tương tự như GA, ethylene có tác dụng đối nghịch với ABA, tuy nhiên vẫn còn thiếu bằng chứng cho thấy việc sản xuất ethylene được tăng cường ở giai đoạn nào khi hạt lúa nảy mầm. Ngoài ra, một số yếu tố di truyền liên quan đến đặc tính giống như khả năng kháng PHS, tính thấm của vỏ hạt, giai đoạn trưởng thành của hạt và các đặc tính nông sinh học khác cũng ảnh hưởng đến quá trình PHS [10].
Chiến lược nghiên cứu và chọn tạo giống cây ngũ cốc kháng lại hiện tượng PHS
Đến nay, có hai chiến lược kháng PHS trên ngũ cốc đã được chứng minh là hiệu quả [5]. Thứ nhất, kiểm soát cơ chế tín hiệu GA kích thích nảy mầm [5]. Nelson và cộng sự (2017) [11] đã tiến hành so sánh mức độ phiên mã toàn hệ gen trên mẫu hạt khô của các dòng đột biến sleepy1-2 so với đối chứng Arabidopsis thaliana nòi sinh thái Landsberg erecta (Ler). Trong đó, gen SLEEPY1 (SLY1) mã hóa cho tiểu phần F-box của E3 ubiquitin ligase SCF, một enzym có vai trò trong phản ứng hình thành phức hợp GID1-GA-DELLA, gây đột biến mất đoạn trên gen SLY1 có thể làm tích lũy quá mức DELLA, từ đó làm giảm khả năng ngủ nghỉ của hạt, vì vậy chúng có thể đóng vai trò kháng PHS [11]. Thứ hai, tăng cường khả năng ngủ nghỉ của hạt thông qua củng cố quá trình sinh tổng hợp và chu trình tín hiệu ABA [5]. Cụ thể, tăng cường biểu hiện của các gen liên quan đến quá trình sinh tổng hợp ABA có thể mang đến tiềm năng tăng cường tính ngủ nghỉ của hạt và kháng PHS [5, 12]. Một số gen có vai trò trong sinh tổng hợp ABA nội sinh ở hạt được điều hòa lúc dịch mã [12]. Ví dụ, một bước quan trọng trong sinh tổng hợp ABA ở thực vật bậc cao là phản ứng phân cắt ôxy hóa cis-epoxycarotenoids dưới sự tham gia của 9-cis-epoxycarotenoid dioxygenase (NCED) [5, 13]. Biểu hiện quá mức gen PtNCED1 kích thích sản sinh ABA nội sinh trong hạt của loài Phaius tankervilliae, từ đó tăng cường khả năng nảy mầm [5, 13]. Trong dị hóa, gen CYP707A mã hóa cho enzym ABA 8′-hydrolase, nhờ đó quá trình hydroxyl hóa của ABA ở vị trí 8' được xảy ra. Những kết quả này đã chỉ ra rằng biểu hiện của gen NCED và CYP707A có thể đóng vai trò quan trọng trong điều hòa ABA nội sinh trong hạt khi ngủ nghỉ [14].
Trong khi đó, các gen khác liên quan đến con đường tín hiệu ABA lại chủ yếu được điều hòa sau dịch mã [12]. ABI5 là một thành viên của họ nhân tố phiên mã bZIP đóng vai trò thiết yếu trong điều hòa quá trình nảy mầm của hạt và sinh trưởng của cây non giai đoạn sớm. Sự cảm nhận tín hiệu ABA ở thực vật chủ yếu phụ thuộc vào các tín hiệu, với hoạt động của thụ thể PYR/PYL/RCAR, yếu tố điều hòa âm tính PPC (phosphatase 2C) và yếu tố điều hòa dương tính SnRK2. Khi không có mặt ABA, PP2C bám vào SnRK2 để loại bỏ gốc phosphate (PO43-), dẫn đến SnRK2 không thể thực hiện quá trình phosphoryl hóa các cơ chất tiếp theo trong chuỗi dẫn truyền tín hiệu. Khi có mặt ABA, thụ thể PYR/PYL/RCAR5 bám vào ABA và tương tác với PP2C để bắt đầu quá trình phosphoryl hóa và gắn gốc phosphate cho SnRK2 để tiếp tục thực hiện phản ứng phosphoryl hóa cho các cơ chất kế sau trong chuỗi dẫn truyền điện tử. ABI5 có chức năng trong cảm nhận tín hiệu ABA thông qua điều hòa sự biểu hiện của các gen mang yếu tố cis- đáp ứng với ABA (ABRE) trên vùng promoter của chúng. Hơn nữa, ABI5 cũng có thể được điều hòa bởi nitric oxide (NO) [12]. Quá trình này cũng mang lại những tiềm năng trong ngăn chặn PHS, đồng thời phục hồi khả năng nảy mầm hạt từ những cây kháng PHS bởi vì NO cũng kích thích sự nảy mầm [5].
Có thể thấy rằng, những kết quả đạt được trong việc tìm hiểu bản chất của cơ chế ngủ nghỉ và nảy mầm ở thực vật về cơ bản đã sẵn sàng cho việc áp dụng vào sản xuất. Tuy nhiên, các cơ chế này ở từng loài ngũ cốc vẫn rất khác nhau. Các đánh giá chất chuyển hóa kết hợp với phân tích hệ protein và hệ phiên mã vẫn cần phải được tiến hành trên giống mẫn cảm và kháng PHS ở các cây ngũ cốc.
Một số kết quả nghiên cứu về kháng nảy mầm sớm trên lúa ở Việt Nam
Những năm gần đây, mưa to bất thường gây ngập úng lại thường rơi đúng vào thời điểm chuẩn bị thu hoạch ở cả hai vụ lúa đông xuân và hè thu tại miền Trung nói chung, tỉnh Thừa Thiên - Huế nói riêng. Nếu đợi chín, lúa sẽ nảy mầm vì ngập úng lâu ngày, còn thu hoạch lúc xanh thì công gặt lúa cao gấp 2 lần so với bình thường, trong khi giá lúa bán ra thị trường giảm 25-30%. Trước thực tế đó, một số nghiên cứu thăm dò đã được nhóm nghiên cứu tại Khoa Công nghệ Sinh học, Học viện Nông nghiệp Việt Nam và Khoa Nông học, Trường Đại học Nông lâm Huế triển khai.
Khảo sát cơ cấu lúa tại tỉnh Thừa Thiên - Huế cho thấy, trong các giống lúa canh tác đại trà tại địa phương hiện nay hiện tượng PHS xảy ra tương đối phổ biến. Đáng chú ý, một số giống lúa bản địa trong vùng, điển hình như Chiêm Tây thể hiện tính kháng PHS rất tốt. Nhóm nghiên cứu đã chọn tạo và phát triển 4 dòng lai trở lại cận giao tái tổ hợp (backcross recombinant-inbred line, BIL) kháng PHS trên nền di truyền của giống Khang Dân 18 (KD18). Trong đó, locus tính trạng số lượng (QTL) qPSR7 đã được xác định nằm gần chỉ thị R34C6 trên nhiễm sắc thể số 7 liên kết chặt với sự phân ly tỷ lệ nảy mầm; trong đó alen từ giống lúa indica TSC3 làm tăng và alen từ giống lúa KD18 làm giảm khả năng kháng nảy mầm trước gặt. Thí nghiệm mô phỏng hiện tượng PHS trong điều kiện nhân tạo (ngâm bông lúa vào chậu nước ở nhiệt độ phòng có chiếu sáng ở cường độ 4000 lux, 12h sáng/12h tối) cho thấy, các dòng lúa cải tiến mang QTL qPSR7 có tỷ lệ nảy mầm thấp (<10%) sau 10 ngày xử lý. Như vậy, có thể thấy rằng, tích hợp QTL qPSR7 đã hạn chế hiện tượng PHS trong điều kiện nhân tạo.
Đánh giá sơ bộ 4 dòng cải tiến BIL mang QTL qPSR7 cho thấy, chúng có đặc tính nông sinh học chính và các yếu tố cấu thành năng suất ở mức tương đương so với giống nền KD18. Trong khảo nghiệm tại Thủy Châu, thị xã Hương Thủy, tỉnh Thừa Thiên - Huế, các dòng có thời gian sinh trưởng ngắn, ít hơn 110 ngày (vụ xuân) và 104-105 ngày (vụ mùa). Cây có dạng hình đẹp, thấp cây (cao 110-115 cm), sức sống tốt, độ dài giai đoạn trỗ và độ cứng cây đều đạt điểm 5, độ thoát cổ bông và độ tàn lá lần lượt đạt điểm 1 và 3. Theo dõi trong hai vụ cho thấy, năng suất thực thu của các dòng kháng PHS ở mức tương đương KD18, đạt khoảng 63,0 tạ/ha (vụ xuân) và 61,7 tạ/ha (vụ mùa). Hiện nay, các dòng lúa đang tiếp tục được triển khai trên đồng ruộng nhằm theo dõi tính ổn định, năng suất thực thu, phản ứng kháng/nhiễm với sâu bệnh hại và chất lượng của cơm. Những kết quả này là tín hiệu tích cực cho việc phát triển các dòng lúa KD18 cải tiến kháng PHS tại những khu vực thường xuyên chịu ảnh hưởng của hiện tượng mưa lớn kéo dài trái mùa.
Thay lời kết
Có thể thấy rằng, Việt Nam nằm trong khu vực chịu ảnh hưởng nặng nề của biến đổi khí hậu. Kiểu thời tiết mưa trái mùa bất thường xảy ra tại các tỉnh miền Trung là điều kiện thuận lợi cho hiện tượng PHS. Mặc dù thiệt hại của PHS đến sản xuất lúa gạo ở miền Trung không quá nặng nề nhưng rõ ràng nó làm giảm chất lượng của lúa gạo nói riêng và các hạt ngũ cốc nói chung. Vì vậy, cải thiện tính kháng PHS cho cơ cấu giống lúa canh tác phổ biến tại các tỉnh miền Trung được xem là một bài toán trong tương lai.
Một số ý tưởng của các nhà khoa học trên thế giới là xử lý hóa chất trên đồng ruộng, như phun ABA, để kiểm soát hiện tượng PHS [12]. Điều này là có thể thực hiện được nhưng không khả thi ở Việt Nam. Chiến lược tăng cường ABA nội sinh trong hạt thông qua chỉnh sửa chính xác các gen sinh tổng hợp và liên quan đến dẫn truyền tín hiệu phụ thuộc ABA vẫn đang được thế giới tập trung nghiên cứu [5]. Vì vậy, việc sử dụng các QTL kháng PHS như qPSR7 đồng thời với phát hiện các gen/QTL khác trong nguồn gen cây lúa bản địa để ứng dụng trong việc cải tiến bộ giống lúa trồng là hướng đi có tính hiệu quả cao và bền vững. Một số kết quả bước đầu của nhóm nghiên cứu trong 123 giống lúa bản địa đã phát hiện 80 giống có tỷ lệ PHS 10 ngày rất thấp (<5,4%). Đây là đặc tính rất quý được giữ lại ở các giống lúa địa phương của Việt Nam cần được nghiên cứu và khai thác.
Tóm lại, để ứng phó với điều kiện thời tiết ngày càng bất thường và khó dự báo, công tác chọn tạo giống lúa cho các tỉnh miền Trung cần tập trung nâng cao khả năng kháng PHS của giống khi bị mưa bão, ngập lụt dài ngày.
TÀI LIỆU THAM KHẢO
[1] E. Kirezci, et al. (2020), "Projections of global - scale extreme sea levels and resulting episodic coastal flooding over the 21st century", Sci. Rep., 10, p.11629.
[2] J.M. Vetch, et al. (2019), "Review: revealing the genetic mechanisms of pre-harvest sprouting in hexaploid wheat (Triticum aestivum L.)", Plant Sci., 281, pp.180-185.
[3] M. Yao, et al. (2020), "Methyltransferase1 and ripening modulate vivipary during tomato fruit development", Plant Physiol., 183, pp.1883-1897.
[4] A. Ali, et al. (2019), "Unraveling molecular and genetic studies of wheat (Triticum aestivum L.) resistance against factors causing pre-harvest sprouting", Agronomy, 9(3), DOI: 10.3390/agronomy9030117.
[5] H. Nonogaki, et al. (2018), "Editorial: seed dormancy, germination, and pre-harvest sprouting", Front. Plant Sci., 9, p.1783.
[6] S. Nakamura (2018), "Grain dormancy genes responsible for preventing pre-harvest sprouting in barley and wheat", Breed Sci., 68, pp.295-304.
[7] P.A. Tuan, et al. (2018), "Molecular mechanisms underlying abscisic acid/gibberellin balance in the control of seed dormancy and germination in cereals", Front Plant Sci., 9, DOI: 10.3389/fpls.2018.00668.
[8] L. Du, et al. (2018), "Endosperm sugar accumulation caused by mutation of PHS8/ISA1 leads to pre-harvest sprouting in rice", Plant J., 95, pp.545-556.
[9] A. Yan, Z. Chen (2020), "The control of seed dormancy and germination by temperature, light and nitrate", Bot. Rev., 86, pp.39-75.
[10] H. Olaerts, C.M. Courtin (2018), "Impact of preharvest sprouting on endogenous hydrolases and technological quality of wheat and bread: a review", Compr. Rev. Food Sci. Food Saf., 17, pp.698-713.
[11] S.K. Nelson, et al. (2017), "Biology in the dry seed: transcriptome changes associated with dry seed dormancy and dormancy loss in the Arabidopsis GA-insensitive sleepy1-2 mutant", Front Plant Sci., 8, DOI: 10.3389/fpls.2017.02158.
[12] M. Nonogaki, H. Nonogaki (2017), "Prevention of preharvest sprouting through hormone engineering and germination recovery by chemical biology", Front Plant Sci., 8, DOI: 10.3389/fpls.2017.00090.
[13] H. Nonogaki (2017), "Seed biology updates - highlights and new discoveries in seed dormancy and germination research", Front Plant Sci., 8, DOI: 10.3389/fpls.2017.00524.
[14] S. Ali, et al. (2020), "Implications of abscisic acid in the drought stress tolerance of plants", Agronomy, 10(9), DOI: 10.3390/agronomy10091323.
Nguyễn Quốc Trung1, Hoàng Minh Chính1, Chu Đức Hà2, Nguyễn Thị Hồng1, Dương Thanh Thủy3, Lê Thị Hiên2, Vũ Minh Thiết4
1Khoa Công nghệ Sinh học, Học viện Nông nghiệp Việt Nam
2Khoa Công nghệ Nông nghiệp, Trường Đại học Công nghệ, Đại học Quốc gia Hà Nội
3Khoa Nông học, Trường Đại học Nông lâm Huế
4Viện Kỹ thuật Công nghệ cao Nguyễn Tất Thành, Trường Đại học Nguyễn Tất Thành
 Về trang trước
Về trang trước Về đầu trang
Về đầu trang